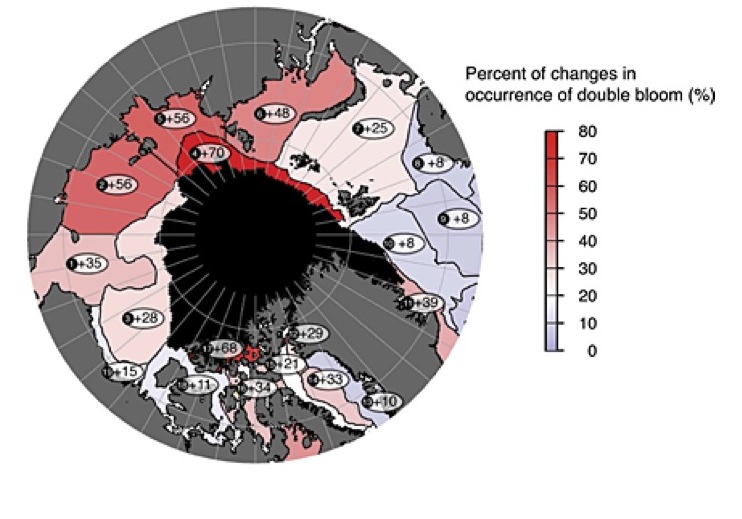
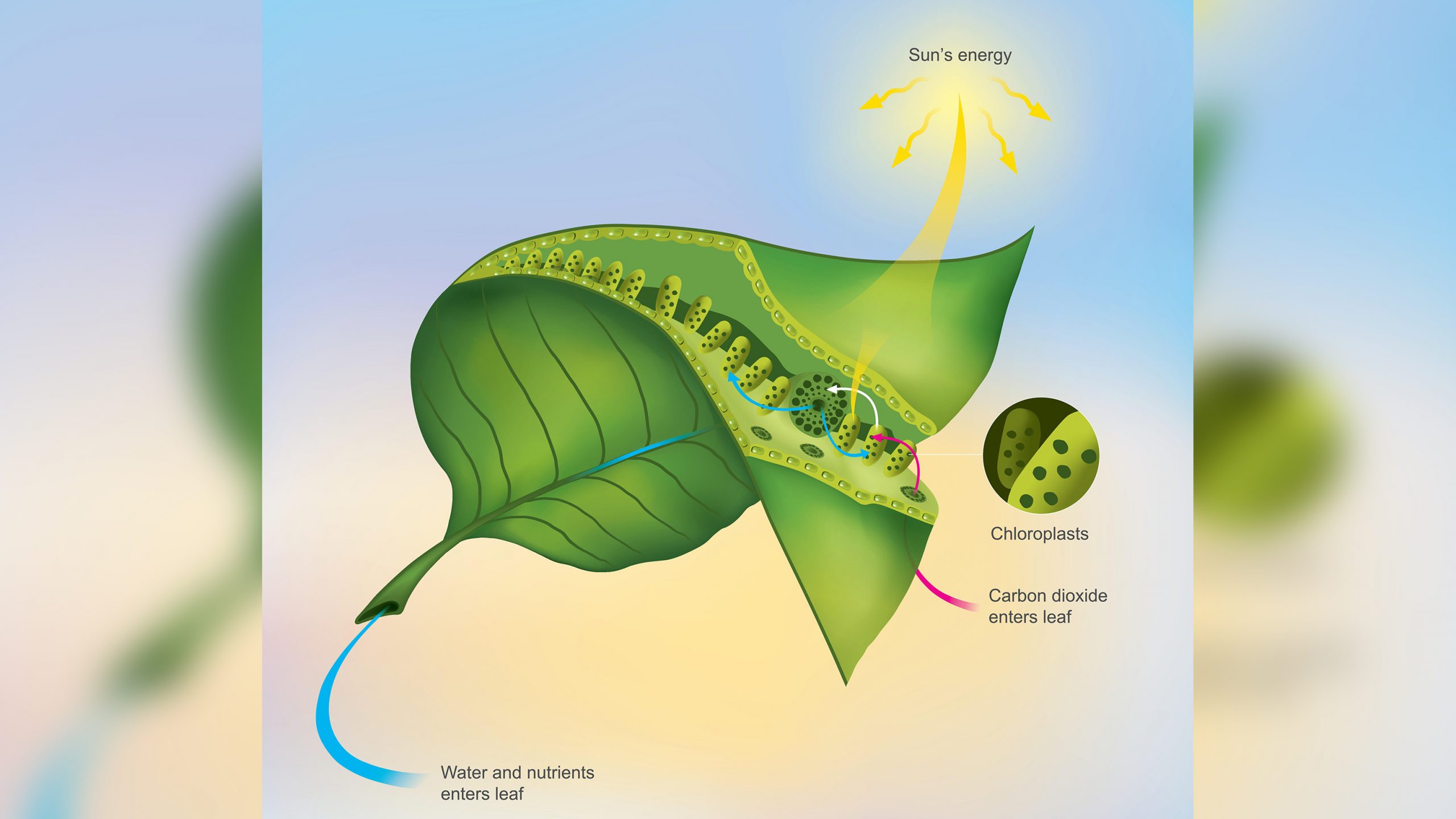
Qu’est-ce que la photosynthèse ?

Les organismes photoautotrophes comme les plantes peuvent transformer des composants minéraux en molécules organiques en utilisant uniquement l’énergie de la lumière solaire. Les chloroplastes sont des organites qui réalisent la photosynthèse. Pour réaliser la photosynthèse, les chloroplastes ont besoin d’un système de membranes très ordonné. Les organismes autotrophes carbonés réalisent la photosynthèse en utilisant des pigments spécifiques pour catalyser deux ensembles distincts de processus.
1. Apprendre la photosynthèse : Un voyage expérimental.
Le rôle du sol et de l’eau. La masse ajoutée d’une plante en croissance provient principalement de la photosynthèse. Dans les plantes, l’eau fournit les électrons utilisés pour réduire le dioxyde de carbone.
Découverte des réactions indépendantes de la lumière. La photosynthèse est un processus en deux étapes. Seule la première étape nécessite directement de la lumière.
Le rôle de la lumière. L’oxygène libéré lors de la photosynthèse des plantes écologiques provient de l’eau, et les atomes de carbone du dioxyde de carbone sont incorporés dans des molécules organiques.
Le rôle de l’énergie réductrice. Les électrons libérés par la division de l’eau réduisent le NADP + ; l’ATP et le NADPH sont ensuite utilisés pour réduire le CO 2 et former des sucres simples.
2. Les pigments captent l’énergie de la lumière solaire.
La biophysique de la lumière. L’énergie de la lumière solaire se présente sous forme de « paquets » appelés photons, qui sont absorbés par les pigments.
Les chlorophylles et les caroténoïdes. Les pigments photosynthétiques absorbent la lumière et récoltent son énergie.
Organisation des pigments en photosystèmes. Un photosystème utilise l’énergie lumineuse pour éjecter un électron excité.
Comment les photosystèmes convertissent la lumière en énergie chimique. Certaines bactéries utilisent un seul photosystème pour produire de l’ATP. Les plantes utilisent deux photosystèmes en série pour générer suffisamment d’énergie pour réduire le NADP+ et produire de l’ATP.
Comment les deux photosystèmes des plantes fonctionnent ensemble. Les photosystèmes II et I conduisent la synthèse de l’ATP et du NADPH nécessaires à la formation de molécules organiques.
3. Les cellules utilisent l’énergie et le pouvoir réducteur captés par les réactions lumineuses pour fabriquer des molécules organiques. Le cycle de Calvin.
L’ATP et le NADPH sont utilisés pour fabriquer des molécules organiques, un processus inversé dans les mitochondries.
Réactions du cycle de Calvin. Le ribulose bisphosphate fixe le CO 2 dans le processus de fixation du carbone.
Photorespiration. L’enzyme qui catalyse la fixation du carbone affecte également la libération de CO 2.
La vie sur terre serait impossible sans la photosynthèse. Chaque atome d’oxygène présent dans l’air que nous respirons faisait autrefois partie d’une molécule d’eau, libérée par la photosynthèse. L’énergie libérée par la combustion du charbon, du bois de chauffage, de l’essence et du gaz naturel, ainsi que par la combustion par notre corps de tous les aliments que nous mangeons – tout cela, directement ou indirectement, a été capturé de la lumière du soleil par photosynthèse. Il est d’une importance vitale que nous comprenions la photosynthèse. La recherche peut nous permettre d’améliorer le rendement des cultures et l’utilisation des terres, des objectifs importants dans un monde de plus en plus surpeuplé. Dans cet article, nous allons examiner la photosynthèse, le processus par lequel les organismes captent l’énergie de la lumière du soleil et l’utilisent pour fabriquer des molécules alimentaires riches en énergie chimique.
Qu’est-ce que la photosynthèse ?
Le chloroplaste, une machine à photosynthétiser
La vie est alimentée par le soleil. L’énergie utilisée par la plupart des cellules vivantes provient en effet du soleil, captée par les plantes, les algues et les bactéries grâce au processus de photosynthèse. La diversité de la vie n’est possible que parce que notre planète est inondée d’énergie provenant du soleil.Chaque jour, l’énergie rayonnante qui atteint la terre équivaut à environ 1 million de bombes atomiques de la taille d’Hiroshima. La photosynthèse capte environ 1 % de cette énorme quantité d’énergie et l’utilise pour fournir l’énergie nécessaire à toute vie.
Le processus de photosynthèse : Résumé
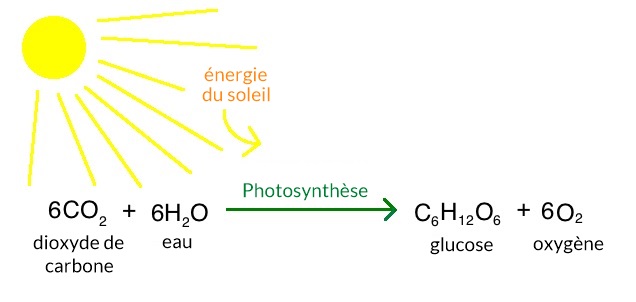
La photosynthèse se produit dans de nombreux types de bactéries et d’algues, ainsi que dans les feuilles et parfois les tiges des plantes écologiques. Les cellules des feuilles des plantes contiennent des organites appelés chloroplastes qui réalisent le processus de photosynthèse. Aucune autre structure dans une cellule végétale n’est capable de réaliser la photosynthèse. La photosynthèse se déroule en trois étapes : (1) la capture de l’énergie de la lumière solaire ; (2) l’utilisation de cette énergie pour produire de l’ATP et du pouvoir réducteur sous la forme d’un composé appelé NADPH ; et (3) l’utilisation de l’ATP et du NADPH pour alimenter la synthèse de molécules organiques à partir du CO 2 présent dans l’air (fixation du carbone). Les deux premières étapes ont lieu en présence de lumière et sont communément appelées les réactions lumineuses. La troisième étape, la formation de molécules organiques à partir du CO 2 atmosphérique, est appelée le cycle de Calvin. Tant que l’ATP et le NADPH sont disponibles, le cycle de Calvin peut se dérouler en l’absence de lumière. L’équation simple suivante résume le processus global de la photosynthèse :
L’intérieur du chloroplaste
Les membranes internes des chloroplastes sont organisées en sacs appelés thylakoïdes, et souvent de nombreux thylakoïdes sont empilés les uns sur les autres en colonnes appelées grana. Les membranes thylakoïdes abritent les pigments photosynthétiques permettant de capter l’énergie lumineuse et les mécanismes de fabrication de l’ATP. Autour du système de membranes thylakoïdes se trouve une substance semi-liquide appelée stroma. Le stroma abrite les enzymes nécessaires à l’assemblage des molécules de carbone. Dans les membranes des thylakoïdes, les pigments photosynthétiques sont regroupés pour former un photosystème.
Chaque molécule de pigment au sein du photosystème est capable de capturer des photons, qui sont des paquets d’énergie. Un réseau de protéines maintient les pigments en contact étroit les uns avec les autres. Lorsque la lumière d’une longueur d’onde appropriée frappe une molécule de pigment dans le photosystème, l’excitation qui en résulte passe d’une molécule de chlorophylle à une autre. L’électron excité n’est pas transféré physiquement – c’est l’énergie qui passe d’une molécule à l’autre. Une analogie grossière de cette forme de transfert d’énergie est la « pause » initiale dans une partie de billard. Si la boule de billard frappe carrément la pointe d’un triangle de 15 boules, les deux boules situées aux extrémités du triangle s’envolent, mais aucune des boules centrales ne bouge. L’énergie passe par les boules centrales pour atteindre les boules les plus éloignées. Finalement, l’énergie arrive à une molécule clé de chlorophylle qui touche une protéine liée à la membrane. L’énergie est transférée sous forme d’électron excité à cette protéine, qui la transmet à une série d’autres protéines membranaires qui utilisent l’énergie pour fabriquer de l’ATP et du NADPH et construire des molécules organiques. Le photosystème agit donc comme une grande antenne, qui recueille la lumière captée par de nombreuses molécules pigmentaires individuelles.
Le rôle du sol et de l’eau
L’histoire de l’apprentissage de la photosynthèse est l’une des plus intéressantes de la science et constitue une bonne introduction à ce processus complexe. L’histoire commence il y a plus de 300 ans, avec une expérience simple mais soigneusement conçue par un médecin belge, Jan Baptista van Helmont (1577-1644). Depuis l’époque grecque, on pense que les plantes tirent leur nourriture du sol, qu’elles aspirent littéralement avec leurs racines. Il a planté un petit saule dans un pot de terre après avoir pesé l’arbre et la terre. L’arbre a grandi dans le pot pendant plusieurs années, au cours desquelles van Helmont n’a ajouté que de l’eau. Au bout de cinq ans, l’arbre était beaucoup plus gros : son poids avait augmenté de 74,4 kilogrammes.
Cependant, toute cette masse supplémentaire ne pouvait pas provenir de la terre, car la terre du pot ne pesait que 57 grammes de moins que cinq ans auparavant ! Avec cette expérience, van Helmont a démontré que la substance de la plante n’était pas produite uniquement à partir de la terre. Il a conclu à tort que l’augmentation de la masse de la plante était principalement due à l’eau qu’il avait ajoutée. Une centaine d’années s’écoulèrent avant que l’histoire ne devienne plus claire. L’indice clé a été fourni par le scientifique anglais Joseph Priestly, dans ses études pionnières sur les propriétés de l’air.
Le 17 août 1771, Priestly a « accidentellement trouvé une méthode pour restaurer l’air qui avait été endommagé par la combustion des bougies ». Il « a mis un brin de menthe [vivante] dans l’air dans lequel une bougie de cire s’était consumée et a constaté que, le 27 du même mois, une autre bougie pouvait être brûlée dans ce même air ». D’une certaine manière, la végétation semblait avoir restauré l’air ! Priestly a constaté que si une souris ne pouvait pas respirer l’air épuisé par la bougie, l’air « restauré » par la végétation n’était « pas du tout gênant pour une souris ». L’indice clé était que la végétation vivante ajoute quelque chose à l’air. Comment la végétation « restaure » l’air ? Vingt-cinq ans plus tard, le médecin néerlandais Jan Ingenhousz a résolu l’énigme. Après plusieurs années de travail, Ingenhousz a reproduit et considérablement étendu les résultats de Priestly, démontrant que l’air n’était rétabli qu’en présence de la lumière du soleil, et uniquement par les feuilles écologiques d’une plante, et non par ses racines. Il a proposé que les parties vertes de la plante effectuent un processus (que nous appelons aujourd’hui photosynthèse) qui utilise la lumière du soleil pour diviser le dioxyde de carbone (CO 2) en carbone et en oxygène. Il a suggéré que l’oxygène était libéré dans l’air sous forme de gaz O 2, tandis que l’atome de carbone se combinait à l’eau pour former des hydrates de carbone. Sa proposition était une bonne supposition, même si la dernière étape a été modifiée par la suite. Les chimistes ont découvert par la suite que les proportions d’atomes de carbone, d’oxygène et d’hydrogène dans les hydrates de carbone sont effectivement d’environ un atome de carbone par molécule d’eau (comme l’indique le terme hydrate de carbone). Un botaniste suisse a découvert en 1804 que l’eau était un réactif nécessaire. A la fin de ce siècle, la réaction globale de la photo-
synthèse pouvait s’écrire comme suit
CO2 + H 2O + énergie lumineuse — → ( CH 2O ) + O2
Il s’avère toutefois qu’il y a plus que cela. Lorsque les chercheurs ont commencé à examiner le processus plus en détail au siècle dernier, le rôle de la lumière s’est avéré d’une complexité inattendue.
Découverte des réactions indépendantes de la lumière
La première équation d’Ingenhousz pour la photosynthèse inclut un facteur que nous n’avons pas abordé : l’énergie lumineuse. Quel rôle la lumière joue-t-elle dans la photosynthèse ? Au début du siècle dernier, le physiologiste anglais F. F. Blackman a commencé à s’intéresser à la question du rôle de la lumière dans la photosynthèse. En 1905, il est arrivé à la conclusion surprenante que la photosynthèse est en fait un processus en deux étapes, dont une seule utilise directement la lumière.
Blackman a mesuré les effets de différentes intensités lumineuses, concentrations de CO 2 et températures sur la photosynthèse. Tant que l’intensité lumineuse était relativement faible, il a constaté que la photosynthèse pouvait être accélérée en augmentant la quantité de lumière, mais pas en augmentant la température.
la quantité de lumière, mais pas en augmentant la température ou la concentration de CO 2. En revanche, à des intensités lumineuses élevées, une augmentation de la température ou de la concentration de CO 2 accélère considérablement la photosynthèse. Blackman a conclu que la photosynthèse se compose d’un premier ensemble de réactions dites « lumineuses », qui sont largement indépendantes de la température, et d’un second ensemble de réactions « sombres », qui semblent être indépendantes de la lumière mais limitées par le CO 2 . Ne vous méprenez pas sur les étiquettes de Blackman : les réactions dites » sombres » se produisent à la lumière (en fait, elles nécessitent les produits des réactions claires) ; leur nom indique simplement que la lumière n’est pas directement impliquée dans ces réactions. Blackman a constaté que l’augmentation de la température accroît la vitesse des réactions sombres de réduction du carbone, mais seulement jusqu’à 35°C environ. Des températures plus élevées entraînent une chute rapide du taux. Comme 35°C est la température à laquelle de nombreuses enzymes végétales commencent à être dénaturées (les liaisons hydrogène qui maintiennent une enzyme dans sa forme catalytique particulière commencent à être rompues), Blackman a conclu que les enzymes doivent effectuer les réactions de l’obscurité.
Le rôle de la lumière
Le rôle de la lumière dans les réactions dites de lumière et d’obscurité a été déterminé dans les années 1930 par C. B. van Niel, alors étudiant diplômé de l’université de Stanford, qui étudiait la photosynthèse chez les bactéries. L’un des types de bactéries qu’il étudiait, la bactérie du soufre pourpre, ne libère pas d’oxygène pendant la photosynthèse ; au lieu de cela, elle transforme le sulfure d’hydrogène (H2S) en globules de soufre élémentaire pur qui s’accumulent à l’intérieur d’elle-même. Le processus que van Niel a observé est le suivant
CO2 + 2 H2S + énergie lumineuse → ( CH2O ) + H2O + 2 S
Le parallèle frappant entre cette équation et celle d’Ingenhousz a conduit van Niel à proposer que le processus généralisé de la photosynthèse soit en fait
CO2 + 2 H2A + énergie lumineuse → ( CH2O ) + H2O + 2 A
Dans cette équation, la substance H 2 A sert de donneur d’électrons. Dans la photosynthèse réalisée par les plantes écologiques, H 2 A est de l’eau, tandis que chez les bactéries sulfureuses pourpres, H 2 A est du sulfure d’hydrogène. Le produit, A, provient de la scission de H 2 A. Par conséquent, l’O 2 produit lors de la photosynthèse des plantes vertes résulte de la scission de l’eau, et non du dioxyde de carbone. Lorsque les isotopes sont devenus d’un usage courant en biologie au début des années 1950, il est devenu possible de tester la proposition révolutionnaire de van Niel. Les chercheurs ont examiné la photosynthèse de plantes écologiques alimentées en eau 18O ; ils ont constaté que le marqueur 18O se retrouvait dans l’oxygène gazeux plutôt que dans les glucides, comme van Niel l’avait prédit :
CO2 + 2 H218 O + énergie lumineuse — → ( CH 2O ) + H2O + 18 O2
Chez les algues et les plantes écologiques, le glucide généralement produit par la photosynthèse est le sucre glucose, qui possède six carbones. L’équation équilibrée complète de la photosynthèse dans ces organismes est donc la suivante
6 CO 2 + 12 H2O + énergie lumineuse — → C6H12 O6 + 6 O 2 + 6 H2O.
Nous savons maintenant que la première étape de la photosynthèse, les réactions lumineuses, utilise l’énergie de la lumière pour réduire le NADP (une molécule porteuse d’électrons) en NADPH et pour fabriquer de l’ATP. Le NADPH et l’ATP de la première étape de la photosynthèse sont ensuite utilisés dans la deuxième étape, le cycle de Calvin, pour réduire le carbone du dioxyde de carbone et former un sucre simple dont le squelette carboné peut être utilisé pour synthétiser d’autres molécules organiques.
Le rôle du pouvoir réducteur
Dans son travail de pionnier sur les réactions lumineuses, van Niel avait également proposé que le pouvoir réducteur (H+) généré par la division de l’eau soit utilisé pour convertir le CO2 en matière organique dans un processus qu’il appelait la fixation du carbone. Avait-il raison ? Dans les années 1950, Robin Hill a démontré que van Niel avait effectivement raison et que l’énergie lumineuse pouvait être utilisée pour générer du pouvoir réducteur. Des chloroplastes isolés de cellules de feuilles étaient capables de réduire un colorant et de libérer de l’oxygène en réponse à la lumière. Des expériences ultérieures ont montré que les électrons libérés par l’eau étaient transférés au NADP + . Arnon et ses collègues ont montré que les chloroplastes éclairés et privés de CO2 accumulent de l’ATP. Si l’on introduit ensuite du CO2, ni l’ATP ni le NADPH ne s’accumulent, et le CO2 est assimilé à des molécules organiques. Ces expériences sont importantes pour trois raisons. Premièrement, elles démontrent fermement que la photosynthèse ne se produit qu’à l’intérieur des chloroplastes. Deuxièmement, elles montrent que les réactions dépendantes de la lumière utilisent l’énergie lumineuse pour réduire le NADP+ et fabriquer de l’ATP. Troisièmement, ils confirment que l’ATP et le NADPH de ce premier stade de la photosynthèse sont ensuite utilisés dans les réactions ultérieures indépendantes de la lumière pour réduire le dioxyde de carbone et former des sucres simples.
Les pigments captent l’énergie de la lumière solaire.
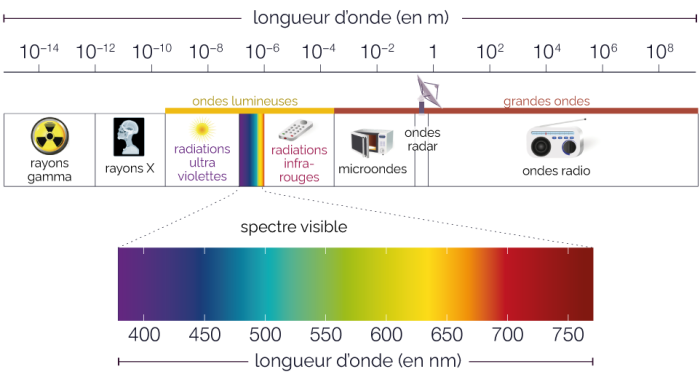
La biophysique de la lumière
Où se trouve l’énergie dans la lumière ? Qu’y a-t-il dans la lumière du soleil qu’une plante peut utiliser pour réduire le dioxyde de carbone ? C’est le mystère de la photosynthèse, le seul facteur fondamentalement différent de processus tels que la respiration. Pour répondre à ces questions, nous devons examiner la nature physique de la lumière elle-même. James Clerk Maxwell avait théorisé que la lumière était une onde électromagnétique, c’est-à-dire qu’elle se déplaçait dans l’air sous forme de champs électriques et magnétiques oscillants. La preuve en a été apportée par une curieuse expérience réalisée dans un laboratoire en Allemagne en 1887. Un jeune physicien, Heinrich Hertz, tentait de vérifier une théorie hautement mathématique qui prédisait l’existence d’ondes électromagnétiques. Pour vérifier l’existence de ces ondes, Hertz a conçu une expérience astucieuse. D’un côté d’une pièce, il a construit un puissant générateur d’étincelles composé de deux grandes sphères métalliques brillantes placées l’une à côté de l’autre sur des tiges hautes et minces. Lorsqu’une charge électrique statique très élevée est accumulée sur une sphère, des étincelles sautent sur l’autre sphère.
Après avoir construit ce dispositif, Hertz entreprend de vérifier si les étincelles créent des ondes électromagnétiques invisibles, appelées ondes radio, comme le prédit la théorie mathématique. De l’autre côté de la pièce, il a placé le premier récepteur radio du monde, un mince cerceau métallique sur un support isolant. Il y avait un petit espace au bas de l’arceau, de sorte que l’arceau ne formait pas tout à fait un cercle complet.
Lorsque Hertz allume le générateur d’étincelles à l’autre bout de la pièce, il voit de petites étincelles traverser l’espace du cercle ! Il s’agit de la première démonstration d’ondes radio. Mais Hertz note un autre phénomène curieux. Lorsque de la lumière UV traverse l’espace du cerceau, les étincelles sont produites plus facilement. Cette facilitation inattendue, appelée effet photoélectrique, a intrigué les chercheurs pendant de nombreuses années. L’effet photoélectrique a finalement été expliqué à l’aide d’un concept proposé par Max Planck en 1901. Planck a développé une équation qui prédit la courbe de rayonnement du corps noir, en partant du principe que la lumière et les autres formes de rayonnement se comportent comme des unités d’énergie appelées photons. En 1905, Albert Einstein a expliqué l’effet photoélectrique en utilisant le concept de photon. La lumière ultraviolette possède des photons d’une énergie suffisante pour que, lorsqu’ils tombent sur la boucle, des électrons soient éjectés de la surface métallique. Les photons avaient transféré leur énergie aux électrons, les faisant littéralement exploser des extrémités du cerceau et facilitant ainsi le passage de l’étincelle électrique induite par les ondes radio. Les longueurs d’onde visibles de la lumière n’ont pas pu éliminer les électrons car leurs photons n’avaient pas assez d’énergie pour libérer les électrons de la surface métallique aux extrémités du cerceau.
L’énergie des photons
Les photons ne possèdent pas tous la même quantité d’énergie. Au contraire, le contenu énergétique d’un photon est inversement proportionnel à la longueur d’onde de la lumière : la lumière à courte longueur d’onde contient des photons plus énergétiques que la lumière à longue longueur d’onde. Les rayons X, qui contiennent beaucoup d’énergie, ont des longueurs d’onde très courtes – beaucoup plus courtes que la lumière visible, ce qui les rend idéaux pour les microscopes à haute résolution. Hertz avait remarqué que la force de l’effet photoélectrique dépendait de la longueur d’onde de la lumière ; les courtes longueurs d’onde sont beaucoup plus efficaces que les grandes pour produire l’effet photoélectrique. La théorie de l’effet photoélectrique d’Einstein fournit une explication : la lumière du soleil contient des photons de nombreux niveaux d’énergie différents, dont seuls certains sont perçus par nos yeux comme de la lumière visible. Les photons les plus énergétiques, à l’extrémité courte du spectre électromagnétique, sont les rayons gamma, dont la longueur d’onde est inférieure à 1 nanomètre ; les photons les moins énergétiques, dont la longueur d’onde peut atteindre des milliers de mètres, sont les ondes radio. Dans la partie visible du spectre, la lumière violette a la longueur d’onde la plus courte et les photons les plus énergétiques, et la lumière rouge a la longueur d’onde la plus longue et les photons les moins énergétiques.
La lumière ultraviolette
La lumière du soleil qui atteint la surface de la terre contient une quantité importante de lumière ultraviolette (UV) qui, en raison de sa longueur d’onde plus courte, possède beaucoup plus d’énergie que la lumière visible. On pense que la lumière UV a été une source d’énergie importante sur la terre primitive lorsque la vie est apparue. L’atmosphère actuelle contient de l’ozone (dérivé de l’oxygène), qui absorbe la plupart des photons UV de la lumière solaire, mais une quantité considérable de lumière UV parvient tout de même à pénétrer dans l’atmosphère. Cette lumière UV est une force puissante qui perturbe les liaisons de l’ADN, provoquant des mutations qui peuvent conduire au cancer de la peau. La perte d’ozone atmosphérique due aux activités humaines menace de provoquer une augmentation considérable de l’incidence des cancers de la peau dans le monde.
Spectres d’absorption et pigments
Comment une molécule « capte » l’énergie de la lumière ? Un photon peut être considéré comme un paquet d’énergie se déplaçant très rapidement. Lorsqu’il frappe une molécule, son énergie est soit perdue sous forme de chaleur, soit absorbée par les électrons de la molécule, ce qui fait passer ces électrons à des niveaux d’énergie plus élevés. L’absorption ou non de l’énergie du photon dépend de la quantité d’énergie qu’il transporte (définie par sa longueur d’onde) et de la nature chimique de la molécule qu’il frappe. Les électrons occupent des niveaux d’énergie distincts dans leurs orbites autour des noyaux atomiques. Pour faire passer un électron à un autre niveau d’énergie, il faut la bonne quantité d’énergie, tout comme pour atteindre le prochain échelon d’une échelle, il faut lever le pied à la bonne distance.
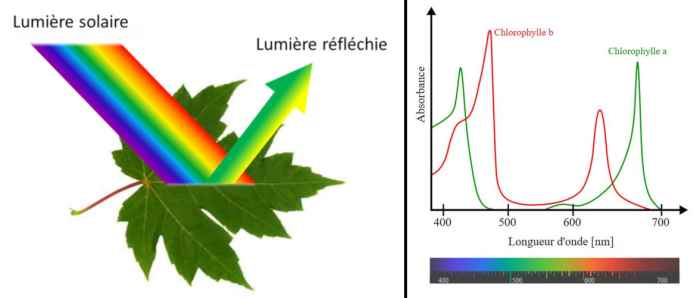
Un atome spécifique ne peut donc absorber que certains photons de lumière, à savoir ceux qui correspondent aux niveaux d’énergie électroniques disponibles de l’atome. Par conséquent, chaque molécule possède un spectre d’absorption caractéristique, c’est-à-dire la gamme et l’efficacité des photons qu’elle est capable d’absorber. Les molécules qui absorbent bien la lumière dans le domaine visible sont appelées pigments. Les organismes ont développé une grande variété de pigments différents, mais seuls deux types généraux sont utilisés dans la photosynthèse des plantes écologiques : les caroténoïdes et les chlorophylles. Les chlorophylles absorbent les photons dans une plage d’énergie étroite. Deux types de chlorophylle dans les plantes, les chlorophylles a et b, absorbent de manière référentielle la lumière violette-bleue et rouge.
Aucun de ces pigments n’absorbe les photons dont la longueur d’onde est comprise entre 500 et 600 nanomètres environ, et la lumière de ces longueurs d’onde est donc réfléchie par les plantes. Lorsque ces photons sont ensuite absorbés par le pigment de nos yeux, nous les percevons comme étant verts. La chlorophylle a est le principal pigment photosynthétique et le seul qui puisse agir directement pour convertir l’énergie lumineuse en énergie chimique. Cependant, la chlorophylle b, agissant comme un pigment accessoire ou secondaire absorbant la lumière, complète et ajoute à l’absorption de lumière de la chlorophylle a.
La chlorophylle b a un spectre d’absorption décalé vers les longueurs d’onde vertes. Par conséquent, la chlorophylle b peut absorber les photons que la chlorophylle a ne peut pas absorber. La chlorophylle b augmente donc considérablement la proportion de photons de la lumière solaire que les plantes peuvent récolter. Un groupe important de pigments accessoires, les caroténoïdes, participent à la photosynthèse en captant l’énergie de la lumière à des longueurs d’onde qui ne sont pas efficacement absorbées par les chlorophylles.
Chlorophylles et caroténoïdes
Les chlorophylles absorbent les photons au moyen d’un processus d’excitation analogue à l’effet photoélectrique. Ces pigments contiennent une structure cyclique complexe, appelée anneau de porphyrine, avec une alternance de liaisons simples et doubles. Au centre de l’anneau se trouve un atome de magnésium. Les photons absorbés par la molécule de pigment excitent les électrons dans l’anneau, qui sont ensuite canalisés par le système de liaisons carbone alternées. Plusieurs petits groupes latéraux fixés à l’extérieur de l’anneau modifient les propriétés d’absorption de la molécule dans différents types de chlorophylle. Le spectre d’absorption précis est également influencé par le microenvironnement local créé par l’association de la chlorophylle avec des protéines spécifiques. Dès qu’Ingenhousz a démontré que seules les parties écologiques des plantes pouvaient « restituer » l’air, les chercheurs ont soupçonné que la chlorophylle était le principal pigment utilisé par les plantes pour absorber la lumière lors de la photosynthèse. Des expériences menées dans les années 1800 ont clairement vérifié ce soupçon. L’une de ces expériences, réalisée par T. W. Englemann en 1882 (figure 10.7), constitue un exemple particulièrement élégant, simple dans sa conception et clair dans son résultat.
Englemann a entrepris de caractériser le spectre d’action de la photosynthèse, c’est-à-dire l’efficacité relative des différentes longueurs d’onde de la lumière pour favoriser la photosynthèse. Il a réalisé toute l’expérience en utilisant une seule lame montée sur un microscope. Pour obtenir différentes longueurs d’onde de lumière, il a placé un prisme sous son microscope, divisant la lumière qui éclaire la lame en un spectre de couleurs. Il a ensuite disposé un filament de cellules d’algues écologiques sur le spectre, de sorte que différentes parties du filament soient éclairées par différentes longueurs d’onde, et a permis aux algues de réaliser la photosynthèse. Pour évaluer la vitesse de la photosynthèse, Englemann a choisi de surveiller le taux de production d’oxygène. Ne disposant pas d’un spectromètre de masse et d’autres instruments modernes, il a ajouté des bactéries aérotactiques (qui recherchent l’oxygène) sur la lame ; il savait qu’elles se rassembleraient le long du filament aux endroits où l’oxygène était produit. Il a constaté que les bactéries s’accumulaient dans les zones éclairées par la lumière rouge et violette, les deux couleurs les plus fortement absorbées par la chlorophylle.
Toutes les plantes, les algues et les cyanobactéries utilisent la chlorophylle a comme pigment primaire. Il est raisonnable de se demander pourquoi ces organismes photosynthétiques n’utilisent pas un pigment comme le rétinal (le pigment de nos yeux), dont le spectre d’absorption est large et couvre la plage des 500 à 600 nanomètres. L’hypothèse la plus probable concerne la photoefficacité. Bien que le rétinal absorbe un large éventail de longueurs d’onde, il le fait avec une efficacité relativement faible. La chlorophylle, en revanche, n’absorbe que deux bandes étroites, mais avec une efficacité élevée. Par conséquent, les plantes et la plupart des autres organismes photosynthétiques atteignent des taux de capture de photons globaux bien plus élevés avec la chlorophylle qu’avec les autres pigments.
Les caroténoïdes sont constitués d’anneaux de carbone liés à des chaînes comportant une alternance de liaisons simples et doubles. Ils peuvent absorber des photons avec une large gamme d’énergies, bien qu’ils ne soient pas toujours très efficaces dans le transfert de cette énergie. Les caroténoïdes participent à la photosynthèse en captant l’énergie de la lumière dont les longueurs d’onde ne sont pas efficacement absorbées par les chlorophylles. Un caroténoïde typique est le ß-carotène, dont les deux anneaux de carbone sont reliés par une chaîne de 18 atomes de carbone avec des liaisons simples et doubles alternées. La division d’une molécule de ß-carotène en deux moitiés égales produit deux molécules de vitamine A. L’oxydation de la vitamine A produit du rétinal, le pigment utilisé dans la vision des vertébrés. Cela explique pourquoi les carottes, qui sont riches en ß-carotène, améliorent la vision.
Organisation des pigments en photosystèmes
Les réactions lumineuses de la photosynthèse se produisent dans des membranes. Chez les bactéries comme celles étudiées par van Niel, la membrane plasmique elle-même est la membrane photosynthétique. Chez les plantes et les algues, en revanche, la photosynthèse est réalisée par des organites qui sont les descendants évolutifs des bactéries photosynthétiques, les chloroplastes – les membranes photosynthétiques existent à l’intérieur des chloroplastes. Les réactions lumineuses se déroulent en quatre étapes :
1. Photo-événement primaire. Un photon de lumière est capturé par un pigment. Le résultat de ce photoévénement primaire est l’excitation d’un électron dans le pigment.
2. Séparation des charges. Cette énergie d’excitation est transférée à un pigment chlorophyllien spécialisé appelé centre de réaction, qui réagit en transférant un électron énergétique à une molécule acceptrice, initiant ainsi le transport d’électrons.
3. Transport d’électrons. L’électron excité est transporté le long d’une série de molécules porteuses d’électrons intégrées à la membrane photosynthétique. Plusieurs d’entre elles réagissent en transportant des protons à travers la membrane, générant un gradient de concentration en protons. Son arrivée à la pompe induit le transport d’un proton à travers la membrane. L’électron est alors transmis à un accepteur.
4. La chimiosmose. Les protons qui s’accumulent d’un côté de la membrane traversent à nouveau la membrane à travers des complexes protéiques spécifiques où la synthèse chimiosmotique de l’ATP a lieu, tout comme dans la respiration aérobie.
Découverte des photosystèmes
Une façon d’étudier comment les pigments absorbent la lumière est de mesurer la dépendance du rendement de la photosynthèse par rapport à l’intensité de l’éclairage, c’est-à-dire la quantité de photosynthèse produite par la quantité de lumière. Lorsque des expériences de ce type sont réalisées sur des plantes, elles montrent que le rendement de la photosynthèse augmente de façon linéaire à faible intensité, mais diminue à des intensités plus élevées, pour finalement saturer à une intensité lumineuse élevée. La saturation se produit parce que toute la capacité d’absorption de la lumière de la plante est utilisée ; une lumière supplémentaire n’augmente pas le rendement car il n’y a rien pour absorber les photons supplémentaires. Il est tentant de penser qu’à la saturation, toutes les molécules de pigments d’une plante sont utilisées.
En 1932, les physiologistes Robert Emerson et William Arnold ont entrepris de vérifier cette hypothèse dans un organisme où ils pouvaient mesurer à la fois le nombre de molécules de chlorophylle et le rendement de la photosynthèse. Dans leur expérience, ils ont mesuré le rendement en oxygène de la photosynthèse lorsque des Chlorella (algues écologiques unicellulaires) étaient exposées à de très brefs flashs lumineux ne durant que quelques microsecondes. En supposant que l’hypothèse de la saturation des pigments était correcte, ils s’attendaient à ce qu’en augmentant l’intensité des flashs, le rendement par flash augmente, jusqu’à ce que chaque molécule de chlorophylle absorbe un photon, qui serait ensuite utilisé dans les réactions lumineuses, produisant une molécule de O 2. De manière inattendue, ce n’est pas ce qui s’est passé. Au contraire, la saturation a été atteinte beaucoup plus tôt, avec seulement une molécule de O 2 pour 2500 molécules de chlorophylle ! Cela a conduit Emerson et Arnold à conclure que la lumière n’est pas absorbée par des molécules de pigment indépendantes, mais plutôt par des groupes de molécules de chlorophylle et de pigments accessoires, que l’on a appelés photosystèmes.
La lumière est absorbée par l’une des centaines de molécules pigmentaires d’un photosystème, qui transfèrent leur énergie d’excitation à une molécule dont le niveau d’énergie est inférieur à celui des autres. Ce centre de réaction du photosystème agit comme un puits d’énergie, en piégeant l’énergie d’excitation. C’est la saturation de ces centres de réaction, et non des molécules individuelles, qui a été observée par Emerson et Arnold.
Architecture d’un photosystème
Dans les chloroplastes et dans toutes les bactéries, sauf les plus primitives, la lumière est captée par de tels photosystèmes. Chaque photosystème est un réseau de molécules de chlorophylle a, de pigments accessoires et de protéines associées, contenus dans une matrice protéique à la surface de la membrane photosynthétique. Comme une loupe qui focalise la lumière sur un point précis, un photosystème canalise l’énergie d’excitation recueillie par n’importe laquelle de ses molécules de pigment vers une molécule spécifique, la chlorophylle du centre de réaction. Cette molécule transmet ensuite l’énergie hors du photosystème afin qu’elle puisse être utilisée pour la synthèse de l’ATP et des molécules organiques. Un photosystème se compose donc de deux éléments étroitement liés :
1. Un complexe d’antenne composé de centaines de molécules de pigments qui recueillent les photons et transmettent l’énergie lumineuse captée au centre de réaction.
2. Un centre de réaction, composé d’une ou plusieurs molécules de chlorophylle a dans une matrice de protéines, qui transmet l’énergie hors du photosystème.
Le complexe d’antenne. Le complexe d’antenne capture les photons de la lumière solaire. Dans les chloroplastes, le complexe d’antenne est un réseau de molécules de chlorophylle liées entre elles et maintenues fermement sur la membrane du thylakoïde par une matrice de protéines. Des quantités variables de pigments accessoires caroténoïdes peuvent également être présentes. La matrice protéique sert en quelque sorte d’échafaudage, maintenant les molécules de pigment individuelles dans des orientations optimales pour le transfert d’énergie. L’énergie d’excitation résultant de l’absorption d’un photon passe d’une molécule de pigment à une molécule adjacente sur son chemin vers le centre de réaction. Après le transfert, l’électron excité dans chaque molécule retourne au niveau de basse énergie qu’il avait avant l’absorption du photon.
Par conséquent, c’est l’énergie, et non les électrons excités eux-mêmes, qui passe d’une molécule de pigment à l’autre.Le complexe d’antenne achemine l’énergie de nombreux électrons vers le centre de réaction.
Le centre de réaction. Le centre de réaction est un complexe protéine-pigment transmembranaire. Dans le centre de réaction des bactéries photosynthétiques violettes, qui est plus simple que dans les chloroplastes mais mieux compris, une paire de molécules de chlorophylle a agit comme un piège pour l’énergie des photons, faisant passer un lectron excité vers un accepteur positionné précisément comme son voisin. Notez qu’ici, c’est l’électron excité lui-même qui est transféré, et pas seulement l’énergie comme nous l’avons vu dans les transferts pigment-pigment. Cela permet à l’excitation du photon de s’éloigner des chlorophylles et constitue la conversion clé de la lumière en énergie chimique. En excitant un électron de la chlorophylle du centre de réaction, la lumière crée un puissant donneur d’électrons là où il n’y en avait pas auparavant. La chlorophylle transfère l’électron excité à l’accepteur primaire, une molécule de quinone, réduisant la quinone et la convertissant en un donneur d’électrons fort. Un donneur d’électrons faible donne alors un électron de faible énergie à la chlorophylle, la ramenant à son état initial. Dans les chloroplastes des plantes, l’eau sert de donneur d’électrons.
Comment les photosystèmes convertissent la lumière en énergie chimique
Les bactéries utilisent un seul photosystème
On pense que les réseaux de pigments photosynthétiques ont évolué il y a plus de 3 milliards d’années dans des bactéries similaires aux bactéries sulfureuses étudiées par van Niel.
1. Electron est joint à un proton pour fabriquer de l’hydrogène. Dans ces bactéries, l’absorption d’un photon de lumière à un pic d’absorption de 870 nanomètres (proche infrarouge, non visible par l’œil humain) par le photosystème entraîne la transmission d’un électron énergétique le long d’une chaîne de transport d’électrons, qui se combine finalement avec un proton pour former un atome d’hydrogène. Chez les bactéries sulfureuses, le proton est extrait du sulfure d’hydrogène, laissant le soufre élémentaire comme sous-produit. Dans les bactéries qui ont évolué plus tard, ainsi que dans les plantes et les algues, le proton provient de l’eau, produisant de l’oxygène comme sous-produit.
2. L’électron est recyclé dans la chlorophylle. L’éjection d’un électron du centre de réaction de la bactérie lui laisse un électron en moins. Avant que le photosystème de la bactérie du soufre puisse fonctionner à nouveau, un électron doit être renvoyé. Ces bactéries ramènent l’électron vers le pigment par un système de transport d’électrons similaire au passage de l’électron dans une pompe à protons qui favorise la synthèse chimiosmotique de l’ATP. Une molécule d’ATP est produite pour trois électrons qui suivent ce chemin. Vu globalement, le trajet de l’électron est donc un cercle. Les chimistes appellent donc le processus de transfert d’électrons menant à la formation d’ATP la photophosphorylation cyclique.
Notez toutefois que l’électron qui a quitté le centre de réaction P 870 était un électron à haute énergie, boosté par l’absorption d’un photon de lumière, alors que l’électron qui revient n’a que l’énergie qu’il avait avant l’absorption du photon. La différence d’énergie de cet électron est le gain de la photosynthèse, l’énergie qui actionne la pompe à protons.
Pendant plus d’un milliard d’années, la photophosphorylation cyclique a été la seule forme de réaction photosynthétique à la lumière utilisée par les organismes. Cependant, sa principale limite est qu’elle n’est orientée que vers la production d’énergie, et non vers la biosynthèse. La plupart des organismes photosynthétiques incorporent le dioxyde de carbone atmosphérique dans les hydrates de carbone. Comme les molécules d’hydrates de carbone sont plus réduites (ont plus d’atomes d’hydrogène) que le dioxyde de carbone, une source de pouvoir réducteur (c’est-à-dire d’hydrogènes) doit être fournie. La photophosphorylation cyclique ne le fait pas. Les atomes d’hydrogène extraits de H 2 S sont utilisés comme source de protons, et ne sont pas disponibles pour se joindre au carbone. Ainsi, les bactéries qui sont limitées à ce processus doivent récupérer les hydrogènes d’autres sources, une entreprise inefficace.
Pourquoi les plantes utilisent deux photosystèmes
Après l’apparition des bactéries sulfureuses, d’autres types de bactéries ont développé une version améliorée du photosystème qui a surmonté les limites de la photophosphorylation cyclique d’une manière simple et efficace : un deuxième photosystème plus puissant utilisant une autre disposition de la chlorophylle a a été combiné
avec l’original. Dans ce deuxième photosystème, appelé photosystème II, les molécules de chlorophylle a sont disposées selon une géométrie différente, de sorte qu’un plus grand nombre de photons de plus courte longueur d’onde et de plus haute énergie sont absorbés que dans le photosystème ancestral, appelé photosystème I. Comme dans le photosystème ancestral, l’énergie est transmise d’une molécule de pigment à une autre dans le complexe d’antennes de ces photosystèmes jusqu’à ce qu’elle atteigne le centre de réaction, une molécule de pigment particulière positionnée près d’un accepteur d’électrons puissant lié à la membrane.
Dans le photosystème II, le pic d’absorption (c’est-à-dire la longueur d’onde de la lumière la plus fortement absorbée) des pigments est d’environ 680 nanomètres ; le pigment du centre de réaction est donc appelé P 680. Chez les plantes, le pic d’absorption des pigments du photosystème I est de 700 nanomètres ; le pigment du centre de réaction est donc appelé P 700. En travaillant ensemble, les deux photosystèmes effectuent un transfert d’électrons non cyclique. Lorsque le taux de photosynthèse est mesuré à l’aide de deux faisceaux lumineux de longueurs d’onde différentes (l’un rouge et l’autre rouge lointain), le taux est supérieur à la somme des taux obtenus avec les faisceaux individuels de lumière rouge et rouge lointain. Ce résultat surprenant, appelé effet de renforcement, peut être expliqué par un mécanisme impliquant deux photosystèmes agissant en série (c’est-à-dire l’un après l’autre), dont l’un absorbe préférentiellement dans le rouge et l’autre dans le rouge lointain.
L’utilisation de deux photosystèmes résout le problème de l’obtention du pouvoir réducteur de manière simple et directe, en exploitant l’énergie de deux photosystèmes. Les électrons proviennent de l’eau, qui retient très fortement ses électrons (potentiel d’oxydoréduction = +820 mV), et aboutissent dans le NADPH, qui retient ses électrons de façon beaucoup plus lâche (potentiel d’oxydoréduction = -320 mV).
Comment les deux photosystèmes des plantes fonctionnent ensemble
Les plantes utilisent les deux photosystèmes évoqués précédemment en série, d’abord l’un puis l’autre, pour produire de l’ATP et du NADPH. Ce processus en deux étapes est appelé photophosphorylation non cyclique, car le trajet des électrons n’est pas circulaire : les électrons éjectés des photosystèmes n’y retournent pas, mais finissent plutôt dans le NADPH. Les photosystèmes sont réapprovisionnés à la place avec des électrons obtenus par la division de l’eau. Le photosystème II agit en premier. Les électrons de haute énergie générés par le photosystème II sont utilisés pour synthétiser l’ATP, puis transmis au photosystème I pour la production de NADPH. Pour chaque paire d’électrons obtenue de l’eau, une molécule de NADPH et un peu plus d’une molécule d’ATP sont produites.
d’une molécule d’ATP sont produites.
Photosystème II
Le centre de réaction du photosystème II, appelé P680, ressemble beaucoup au centre de réaction des bactéries pourpres. Il est constitué de plus de 10 sous-unités protéiques transmembranaires. Le complexe d’antenne capteur de lumière est constitué de quelque 250 molécules de chlorophylle a et de pigments accessoires liés à plusieurs chaînes de protéines. Dans le photosystème II, les atomes d’oxygène de deux molécules d’eau se lient à un groupe d’atomes de manganèse qui sont intégrés dans une enzyme et liés au centre de réaction. D’une manière mal comprise, cette enzyme divise l’eau, en retirant les électrons un par un pour remplir les trous laissés dans le centre de réaction par le départ des électrons excités par la lumière. Dès que quatre électrons ont été retirés des deux molécules d’eau, l’O2 est libéré.
Le chemin vers le photosystème I
Le principal accepteur d’électrons pour les électrons excités par la lumière qui quittent le photosystème II est une molécule de quinone, comme c’était le cas dans le photosystème bactérien décrit précédemment. La quinone réduite qui en résulte (plastoquinone, symbolisée par Q) est un puissant donneur d’électrons ; elle transmet l’électron excité à une pompe à protons appelée complexe b6-f, intégrée à la membrane thylakoïde. L’arrivée de l’électron énergétique amène le complexe b6-f à pomper un proton dans l’espace thylakoïde. Une petite protéine contenant du cuivre appelée plastocyanine (symbolisée par pC) transporte alors l’électron vers le photosystème I.
Fabrication de l’ATP : Chemiosmose
Chaque thylakoïde est un compartiment fermé dans lequel les protons sont pompés du stroma par le complexe b 6 -f. La division de l’eau produit également des protons supplémentaires qui contribuent au gradient. La membrane thylakoïde étant imperméable aux protons, ces derniers repartent presque exclusivement par les canaux fournis par les ATP synthases. Ces canaux font saillie comme des boutons sur la surface externe de la membrane thylakoïde. Lorsque les protons sortent du thylakoïde par le canal de l’ATP synthase, l’ADP est phosphorylé en ATP et libéré dans le stroma, la matrice fluide à l’intérieur du chloroplaste. Le stroma contient les enzymes qui catalysent les réactions de fixation du carbone.
Photosystème I
Le centre de réaction du photosystème I, appelé P700, est un complexe transmembranaire composé d’au moins 13 sous-unités protéiques. L’énergie lui est fournie par un complexe d’antenne composé de 130 molécules de chlorophylle a et de pigments accessoires. Le photosystème I accepte un électron de la plastocyanine dans le trou créé par la sortie d’un électron excité par la lumière. Cet électron qui arrive n’a absolument pas perdu toute son énergie excitée par la lumière ; il en reste presque la moitié. Ainsi, l’absorption d’un photon d’énergie lumineuse par le photosystème I porte l’électron quittant le centre de réaction à un niveau d’énergie très élevé. Contrairement au photosystème II
et le photosystème bactérien, le photosystème I ne dépend pas des quinones en tant que
accepteurs d’électrons. Au lieu de cela, il transmet les électrons à une protéine fer-soufre
appelée ferrédoxine (Fd).
Fabrication du NADPH
Le photosystème I transmet des électrons à la ferrédoxine sur le côté stromal de la membrane (à l’extérieur du thylakoïde). La ferrédoxine réduite porte un électron à très haut potentiel. Deux d’entre eux, provenant de deux molécules de ferrédoxine réduite, sont ensuite donnés à une molécule de NADP + pour former du NADPH. La réaction est catalysée par la NADP réductase, une enzyme liée à la membrane. Étant donné que la réaction se produit du côté stromal de la membrane et qu’elle implique l’absorption d’un proton pour former le NADPH, elle contribue davantage au gradient de proton établi pendant le transport photosynthétique des électrons.
Fabriquer plus d’ATP
Le passage d’un électron de l’eau au NADPH dans la photophosphorylation non cyclique décrite précédemment génère une molécule de NADPH et un peu plus d’une molécule d’ATP. Cependant, la construction de molécules organiques nécessite plus d’énergie que cela – il faut une molécule et demie d’ATP par molécule de NADPH pour fixer le carbone. Pour produire l’ATP supplémentaire, de nombreuses espèces végétales sont capables de court-circuiter le photosystème I, en faisant passer la photosynthèse en mode de photophosphorylation cyclique, de sorte que l’électron excité par la lumière qui quitte le photosystème I est utilisé pour produire de l’ATP au lieu de NADPH. L’électron énergétique est simplement renvoyé au complexe b 6 -f au lieu de passer au NADP+. Le complexe b6-f expulse un proton, qui s’ajoute au gradient de protons à l’origine de la synthèse chimiosmotique de l’ATP. Les proportions relatives de photophosphorylation cyclique et non cyclique dans ces plantes déterminent les quantités relatives d’ATP et de NADPH disponibles pour la construction de molécules organiques.